1. Introduction
Kombucha is a traditional beverage usually obtained from the fermentation of black or green tea (sweetened with 5–8% of sugar) by a symbiotic microbial consortium, which is mainly composed of acetic acid bacteria (AAB) and osmophilic yeasts [
1]. The peculiarity is that the microorganisms are embedded in a cellulose floating matrix that is produced by AAB. Recently, high-throughput sequencing (HTS) has deeply investigated the relative abundance of the microbial community that is involved in the fermentation of green and black teas [
2,
3]. In particular, authors reported the dominant presence of
Komagataeibacter spp. and
Acetobacter spp. and less than 1% of
Lactobacillus spp., among bacteria;
Zygosaccharomyces spp. and
Brettanomyces spp. are the most abundant yeasts [
2]. Kombucha is known for its nutraceutical properties and it has been consumed in Asia for thousands of years, with its preparation dating back approximately to 220 B.C. [
4]; in the last ten years, the market interest has moved to the whole world [
5], and also home brewing is now a common practice. Substrates, other than black and green teas, can also be used, such as Jerusalem artichoke tuber extracts, wine, milk, fruit juices, and plant infusions [
6], bringing nutritional and health benefits that are based on the selected cultivation medium [
7,
8,
9]. Kombucha analysis is mainly focused on the chemical compounds which confer to the fermented beverage its nutritional and antioxidant value. In particular, at the end of the fermentation process (10–20 days) at a temperature in the range of 20–30 °C, Kombucha is rich in organic acids (acetic, glucuronic, gluconic acids), vitamins and tea polyphenols, and the low pH avoid bacterial contamination [
6]. The beverage has a refreshing taste and consumers have the feeling that Kombucha is beneficial and improve digestion with a regular consumption [
10]. Green and black tea have received considerable attention in recent years as functional beverages due to the high amount of functional compounds, such as polyphenols, flavonoids, and saponins [
11]; polyphenols have been widely studied also in kombucha and they basically increase along the fermentation process [
12,
13,
14]. The fermentation length, temperature, tea quality, sucrose concentration, and the fermenting microbial consortia strictly influence the chemical and anti-oxidant properties of the final beverage [
6,
10]. In particular, tea quality and composition varies with the species, season, age of the leaves (plucking position), climate, and horticultural practices [
15,
16], thus immediately influencing the polyphenols composition of teas and consequently of the fermented product. Moreover, the positive evidence in human health is difficult to show because of the difficulty to organize randomised and controlled intervention trials, which are also very expensive. However, several authors reported a wide spectrum of beneficial activities, studies in vitro or in animal models, including antimicrobial activity against foodborne and human pathogens, hepatic detoxification in rats, anti-inflammatory, hypocholesterolemic, anti-proliferative, and hypoglycemic activities [
6,
17]. To date, only one study has been applied in human subjects research, examining the health benefits of kombucha [
18].
In this work, kombucha was prepared from a consortium of symbiotic microorganisms using two traditional substrates, green and black teas, and a never tested substrate, the tea obtained from rooibos (
Aspalathus linearis) leaves. Microbial diversity was analysed during fermentation both in the cellulolytic pellicles and in the corresponding kombuchas, using culture-dependent methods and Illumina high-throughput sequencing. Additionally, total polyphenols, flavonoids, and anti-oxidant activities were monitored and substrate and metabolite concentrations were quantified. We also investigated whether the kombucha from rooibos exerts a possible protection/restoring effect in mouse fibroblast cell lines against the oxidative stress induced by H
2O
2. This work provides a deep insight on traditionally prepared Kombuchas and gains new knowledge on the use of rooibos herbal tea, which has never been considered as a substrate, in spite of its well-known bioactivity and potential health benefits [
19,
20].
4. Discussion
Fermented beverages are more and more consumed as consumers see the fermentation as a gentle method of preservation, improving well-being and reducing the risk of disease. Consumers of kombucha are increasing worldwide, the brew being a delicious combination of a delicate sour taste, antioxidant properties of tea extracts, and the potentially beneficial effect of fermenting bacteria. Kombucha prepared from black and green teas have been widely studied, although in depth microbiological studies on both bacteria and yeasts with a combination of culture dependent and molecular techniques have not been performed up to now. On the contrary, to the best of our knowledge, kombucha prepared from rooibos has not been studied yet. Rooibos tea was selected as a substrate, because of its reported beneficial action against human diseases [
49,
50], thus hypothesizing a possible additional nutraceutical value beyond the fermentation process. In addition to the focus on a new substrate, this study examines the influence of the different tea substrates on the microbial composition and the presence of antioxidant molecules.
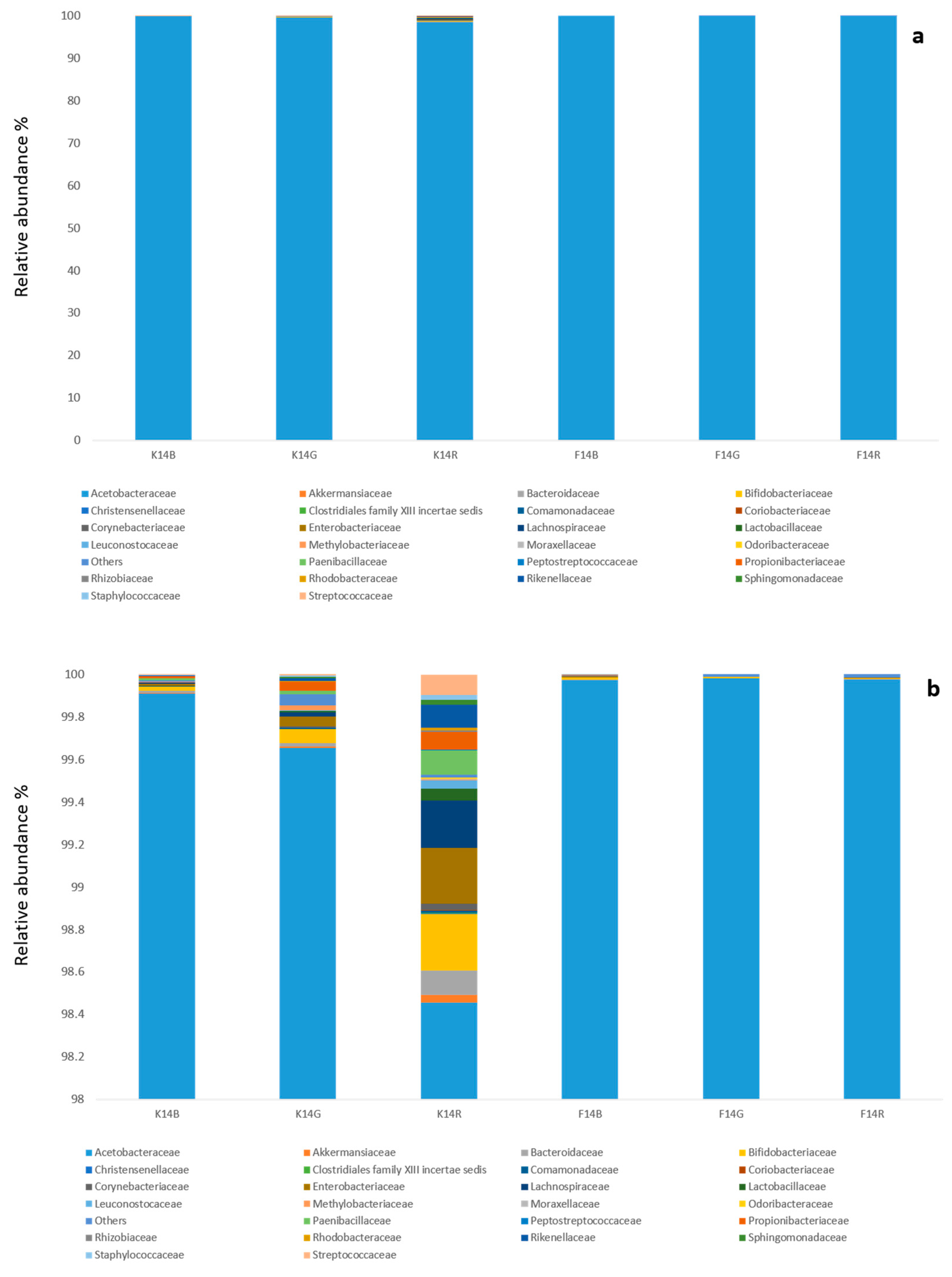
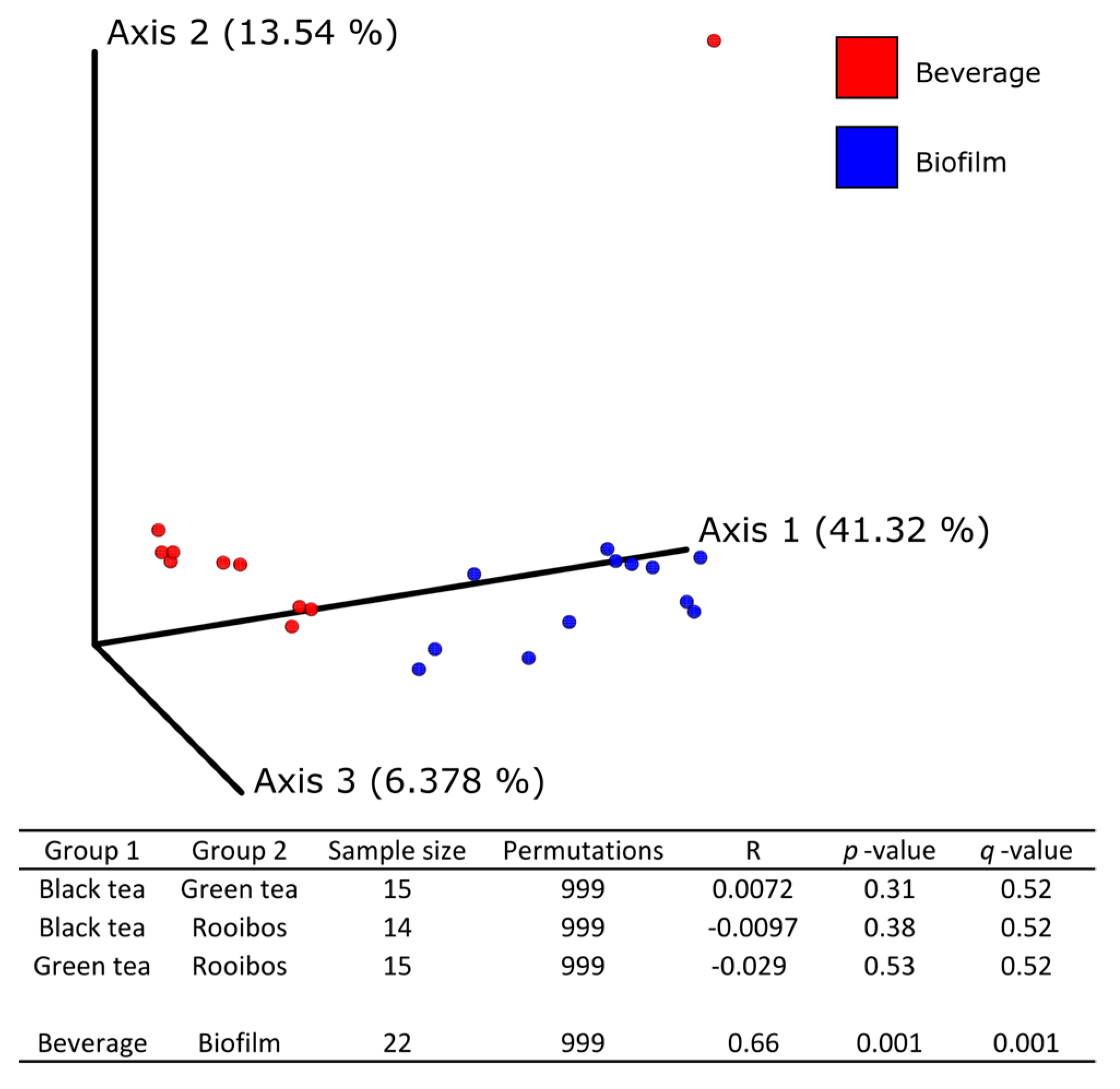
The combination of culture-dependent and culture-independent techniques is a useful approach to mitigating the main problems that arise from culturing (e.g., low-throughput, sensitivity of selective media, slow growing microorganisms, temperature sensitivity); in addition, it allows a deeper insight into microbial diversity and the isolation of some strains of biotechnological interest. Both culturing and HTS showed the dominance of
Komagataeibacter spp. in all biofilms and kombuchas, and HTS conferred this abundance to
K. saccharivorans with no differences among tea types. This is in accordance with other published studies [
3], which mainly found the same species in green and black Kombucha fermented at 30 °C, a temperature that is comparable to that used in this study. Overall, most of the authors performing culture-independent analysis are in substantial agreement with these findings at the genus level [
2,
51], probably determined by a selection process due to the harsh acidic environment at the end of the fermentation process (pH 2.8, as detected in our kombuchas). However, differences can be detected at the species level, based on the assumption that the starter cultures and fermentation temperature may confer the microbiological connotation to the final beverage [
2,
3].
Culture-dependent techniques allowed for the isolation of a limited number of AAB identified as
K. intermedius and
K. rhaeticus, which have been previously identified in fruit juices [
52]. These species have notably high ethanol and acetic acid tolerance, they are GlcUA producers, and they produce cellulose at a high rate, [
53,
54]. The species
K. intermedius has been used in a wide range of food, industrial, and pharmaceutical applications, since not only it has a high ethanol and acetic acid tolerance but it is also an effective producer of exopolysaccharides, nanofibrillated cellulose, and GlcUA [
55]. Indeed, it was mainly isolated from KB and KR, which possess the highest GlcUA concentrations.
Gluconobacter entanii, mainly isolated from KR, is a species originally found in a submerged spirit vinegar fermentation factory [
56], as this species needs high acetic acid concentration and does not produce cellulose. However, due to the enrichment method, it was not possible to get a reliable percentage of their presence in the three substrates. Nevertheless, when considering the decrease in sucrose concentration and the significant increase of acetic acid during the fermentation in all substrates and the consequent GlcUA production, the AAB populations seemed to be consolidated in the beverages without any competition. Although the relative abundance of Firmicutes and Actinobacteria was low (less than 0.3%), a small community belonging to these phyla was detected, as in Marsh et al. [
2] and De Filippis et al. [
3]; members of both phyla were not detectable in the biofilm, but only in the fermented beverage, in particular in KR, where the lowest AA and EtOH concentrations were measured.
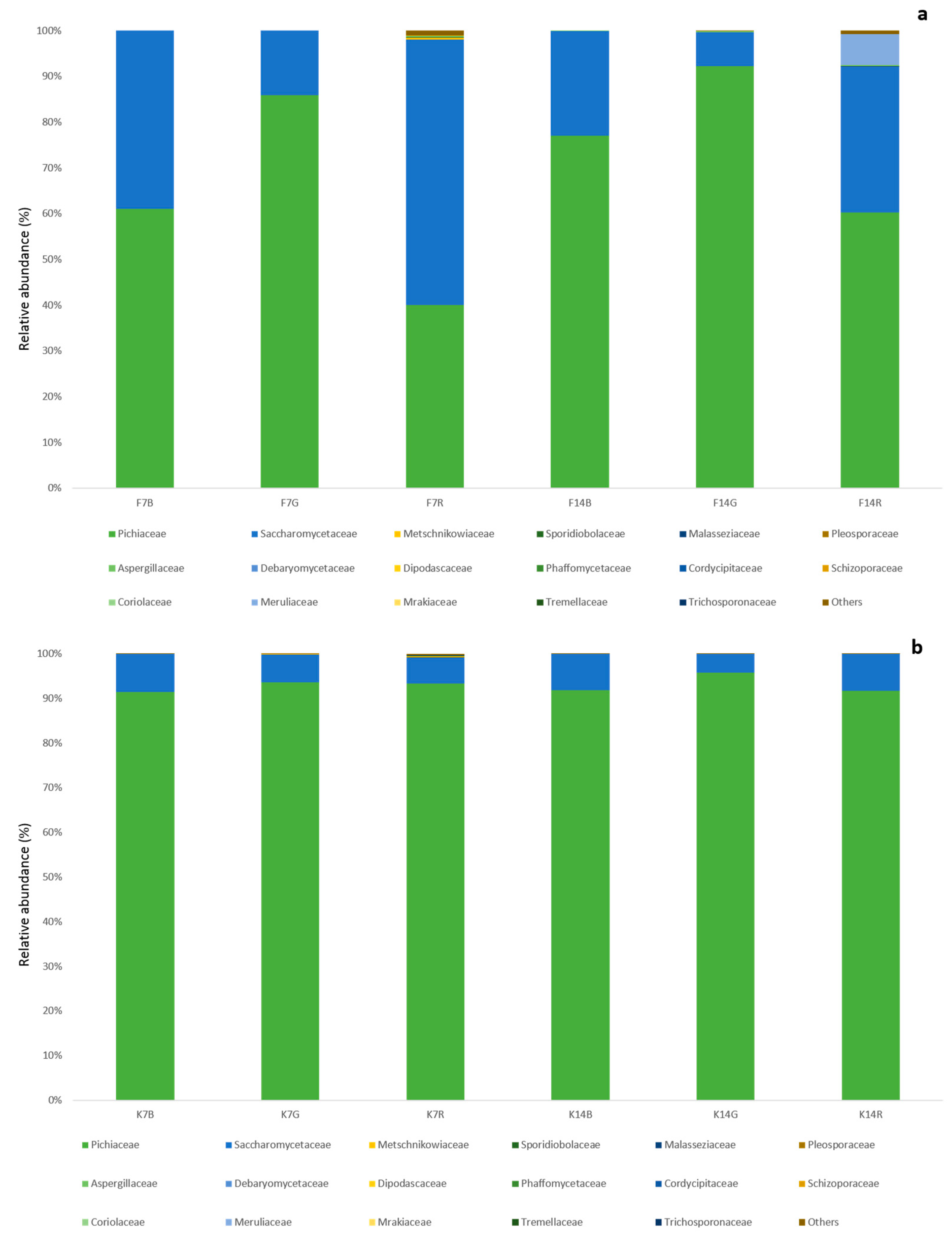
The yeast population that was detected in the different studies in the literature is heterogeneous [
2,
51], indicating different dominant genera in the different studies. Differently from Marsh et al. [
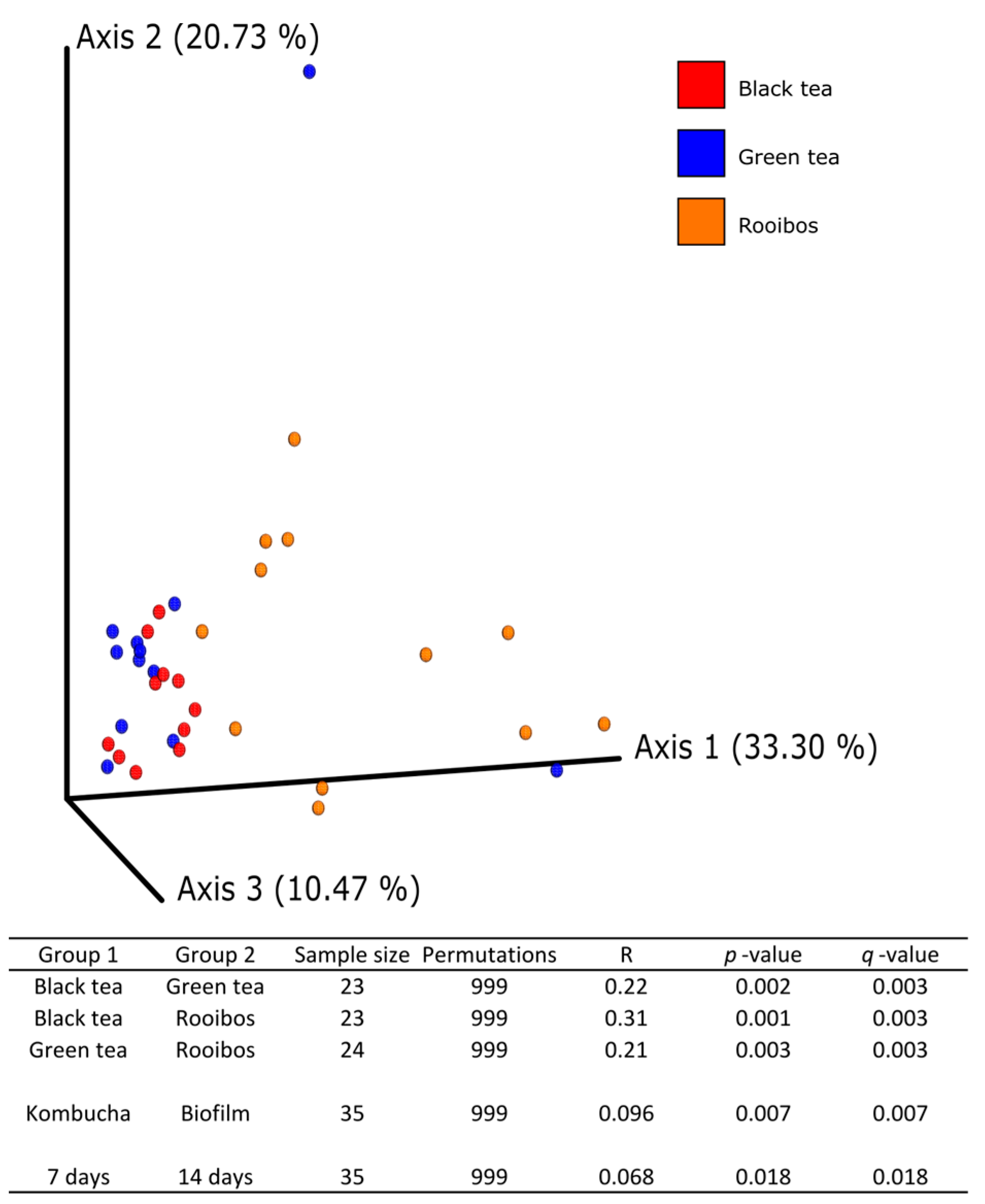
2], the relative abundance of yeasts in the fermented teas did not match that of the biofilm, thereby suggesting that the fungal composition of the cellulosic pellicle used to inoculate the teas is subjected to a selection process during the fermentation, depending on the substrate as well as physical and chemical changes in the medium. Moreover, it appears that the choice of tea substrate has an increasing effect on fungal alpha diversity of kombucha as the fermentation time increases, while the difference in fungal community diversity between kombucha and biofilm decreases.
The two dominant genera found in this study are typical of kombucha [
2,
53,
57]; in particular,
Brettanomyces (the anamorph of
Dekkera) has adapted to harsh and limiting environmental conditions, with low pH values [
58,
59], and it is also often associated with high-ethanol biotechnological habitats [
60,
61]. Interestingly, Nguyen et al. [
53] selected a co-culture of a
B. bruxellensis strain (KN89) and a
K. intermedius strain (KN89) for optimal GlcUA production. Although the relative abundance of
B. bruxellensis in our experiments is low, isolation by plating allowed for the identification of only this species within the
Brettanomyces genus, thus hypothesizing that the unidentified
Dekkera spp. by HTS could be ascribed to the same species.
Chemical analysis showed that the concentrations of organic acids and monosaccharides increased during the fermentation, as already shown by other studies [
12,
13,
62]. Nevertheless, as reviewed by Jayabalan et al. [
6], chemicals are strictly correlated with many parameters (temperature, fermentation periods, pellicle origin, etc.). The different detected concentrations may be correlated with the abundance of acid producing strains as already discussed above. The highest acetic acid and GlcUA concentrations were found in KB as indicated in other published studies [
13,
62]. Concerning KR, any reference value is available, being the rooibos kombucha analysed for the first time in the present work; however, the increasing concentration of GlcUA with time and the low concentration of acetic acid could be considered interesting properties due to the detoxifying properties of GlcUA [
63,
64], and the more delicate taste that was conferred by the lower acetic acid amount. Moreover, GlcUA increases the polyphenols bioavailability [
65]. With respect to antioxidant activity and cathechins, the results are in accordance with authors, stating that cathechin compounds are degraded by bacterial and yeast activity in simpler molecules, thus increasing the antioxidant power [
13,
66,
67]. This aspect is extremely important in a fermented beverage, since antioxidant molecules of teas and plant extracts have scavenger action against free radicals, thus possibly protecting against oxidative damage. Indeed, several studies have already investigated the biological activities of Kombucha prepared from traditional substrates in in vitro cell models and animal models [
20,
68,
69,
70]. Contrary to KB and KG, KR does not contain catechins, since rooibos is not a source of cathechins and this may be the cause of the lower antioxidant activity shown by the DPPH and FRAP assays. However, it contains other molecules, including aspalathin, isoorientin, orientin, and rutin (mainly), and, at lower concentrations, iso-vitexin, vitexin, isoquercitrin and hyperoxide, quercetin, luteolin, and chrysoeryol, all with antioxidant activity [
71]. It has been shown that the aspalathin and nothofagin could be potential therapeutic agents for the treatment of various severe vascular inflammatory diseases via the inhibition of the HMGB1 signaling pathway [
72]. Sanderson et al. [
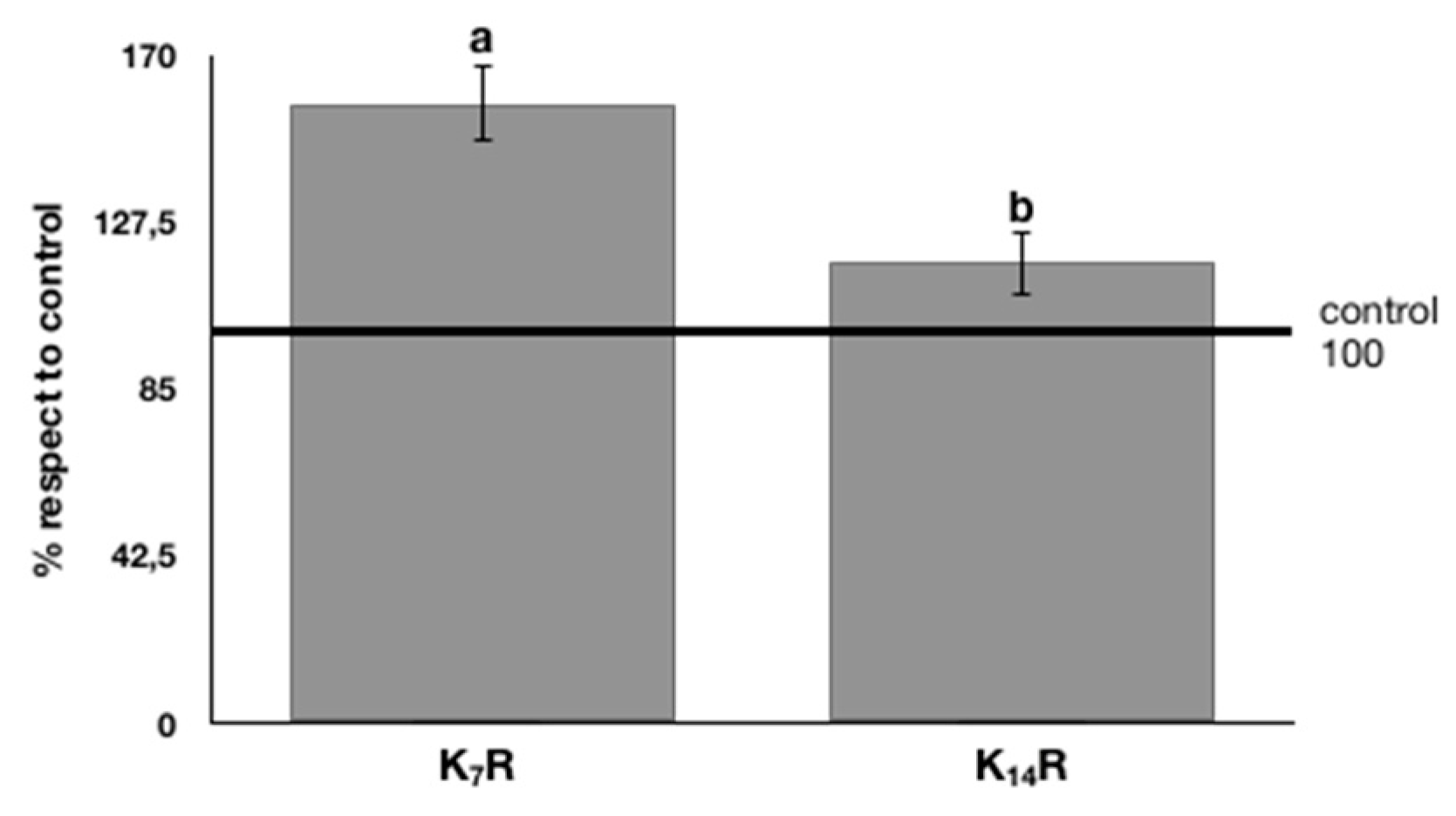
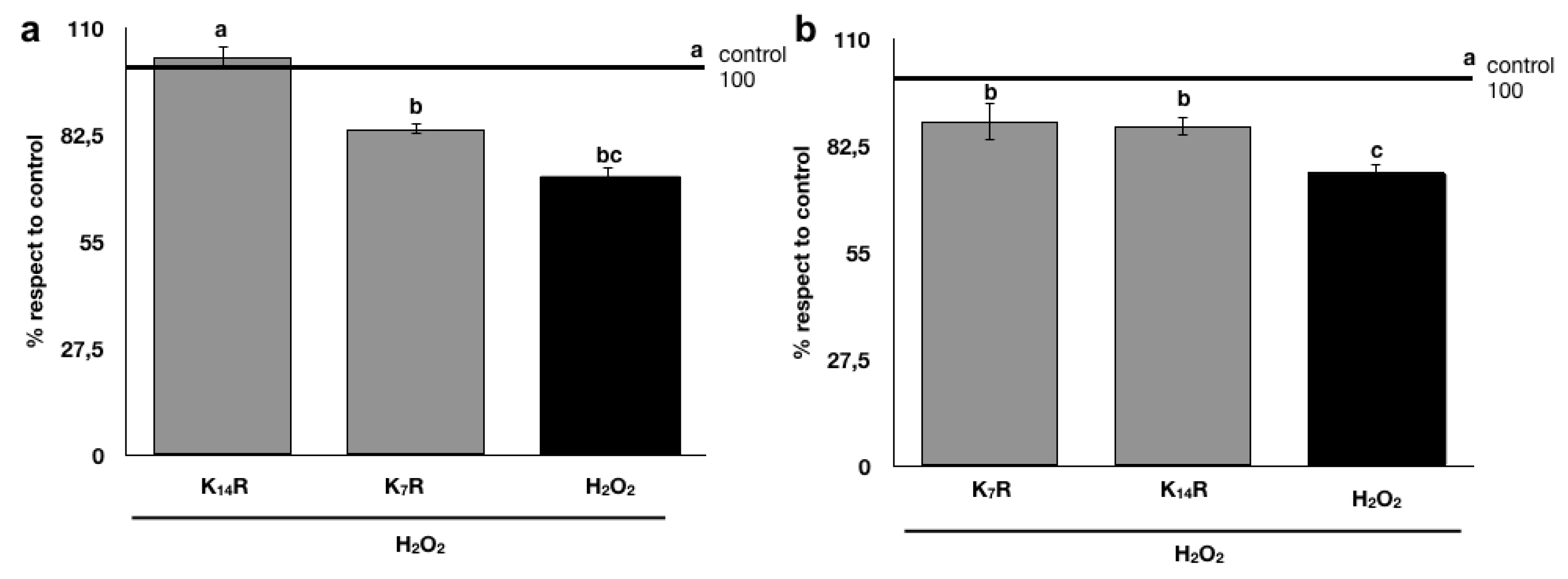
73] showed that hot water-soluble solids from rooibos inhibited adipogenesis and affected adipocyte metabolism, suggesting its potential role in preventing obesity. These observations, therefore, supported the choice of a deeper investigation on the health benefit of rooibos fermented beverage, although the measured antioxidant activity could not be attractive. To the best of our knowledge, this is the first study investigating the possible beneficial activity of KR against the oxidative stress using murine cell models. In the present paper, significantly positive effects on limiting the oxidative stress in the adopted cell model (fibroblast, oxidation induced by H
2O
2 treatment before and after kombucha treatments) was observed for rooibos kombucha. The protective effect of rooibos kombucha on fibroblasts was found to be in line with Pringle et al. [
74], who showed that rooibos infusion decreased cellular oxidative stress and attenuated apoptotic/necrotic cell death in human dermal fibroblasts, by using an in vitro model to mimic diabetic wounds. These results highlighted some properties that support the potential therapeutic action of rooibos and its application in wound healing.


 ,
, 









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}